Mitosis, Cytokinesis and the Cytoskeleton CS6
When observed under time-lapse
photography, the rounding up of the cell, the break up of the nuclear membrane,
the nuclear condensation of the “line-dance” of the chromosomes, and the
eventual splitting of the cells seems directed by an unseen intelligence. Yet of course the whole program is scripted
by genes. Most animal cells divide by
forming a constriction ring that progressively tightens to pinch the daughter
cells apart; plant cells, fungi and fission yeast form a septum between the two
new cells. Budding yeast do something in between by producing a bud off the
parent cell that expands and is then pinched off. The cytoskeleton has a very
large part to play in all of this, and it is likely that the original purpose
in having a cytoskeleton was to direct the distribution of genomes at mitosis.
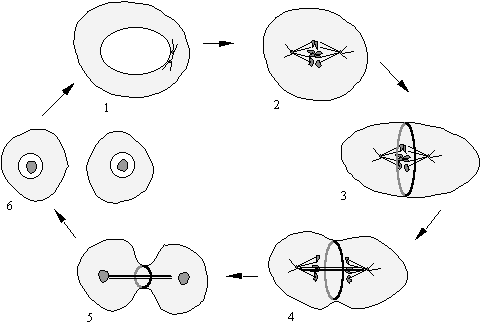
Figure 37. The Cell
Cycle. 1.The centrosome
divides. 2.The nuclear envelope breaks down, chromosomes condense, spindle
poles form. 3. Cleavage furrow (contractile ring) assembles. 4. Cleavage furrow
contracts as chromosomes move off towards the poles. 5. Mid-body forms as the
cleavage furrow narrows further, the chromosomes de-condense and the nuclear
envelope reforms. 6. Cytokinesis is now complete, sometimes the mid-body is
left behind. Each daughter cell grows in size to complete the cycle.
The Centrosome
The centrosome (or microtubule organising
centre, MTOC) is so called because it usually occupies the centre of the cell
or close by. It is a very complex
organelle, capable of replication. The
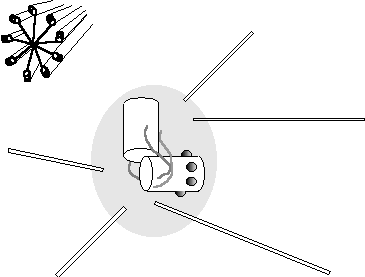
structure of the centrosome is not yet fully characterised (Figure 38). Two short bundles (centrioles) containing
odd triple microtubule-like arrays are evident, these are at approximately 90o
w.r.t. each other. Vague filamentous
material connects these

Microtubule Pericentriolar material Centrioles
Figure 38


![]()
.
Figure 39.
Centrosome duplication.
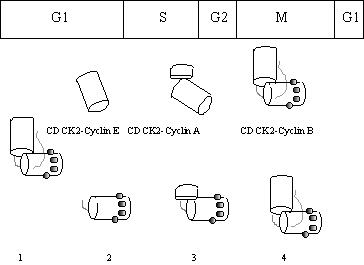
CdcK2-cyclin E is required for centrosome duplication that
starts in G1 (2). In the S phase each centriole grows a
daughter centriole close to its base at about 90O to it (3). This is complete by G2 and two
complete centrosomes are formed by growth of the accessory protein structures
such as the pericentriolar region and filaments. At M phase, (4) the
two centrosomes move apart to occupy opposite ends of the spindle. Cytoskinesis then follows bringing the now
single centrosome back to the start of the cycle (1).
Cdk2-cyclin E activity is required for the splitting of the
two centrioles, these then form nuclei onto which the other components add at
subsequent stages. The other cyclins
active at the other stages are shown, but it is not yet known how or even if,
these co-ordinate centrosome duplication.
Under certain circumstances, centrosomes can originate spontaneously in
the cytoplasm, so there no absolute requirement for pre-existing centrioles. This is unlike most organelles such as
mitochondrial where a cell can only grow new mitochondria from an old one. The centrosome is only half way from being
an organelle and a protein complex.
Microtubule-based motors co-operate to form,
maintain and regulate bipolar spindles.
In addition to
binding membranes, many microtubule motors also bind other microtubules with
the non-motor end and so can bind two microtubules. Minus end directed motors such as dyneins can therefore gather
minus ends together (Figure 40a). These
simple “rules” if applied at the correct time give rise to fairly complex
geometries.
A Likewise, plus end directed
motors (Figure40b) can sort out microtubules arrays bound at their minus
ends by dyneins into anti-parallel arrays:-
B
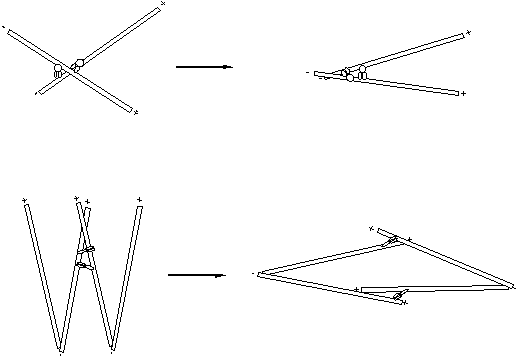
Figure 40
B |
The
position of the cleavage furrow is determined by anti-parallel microtubule
arrays (usually the spindle). Classic
experiments by Rappaport demonstrated the importance of microtubule arrays in
the determination of the position of the cleavage furrow.
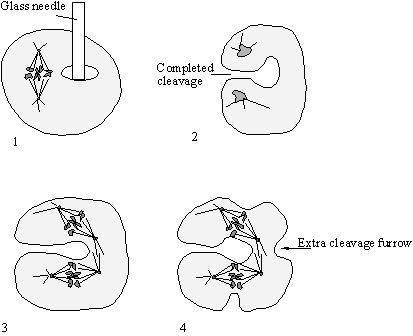
Figure 41. 1. A glass needle was used to create a hole in a cell that was
about to divide. 2. A cleavage furrow appeared between the separated chromosome
masses as usual but the cell did not separate because of its shape. 3. At the
subsequent round of division spindle poles were created at the two chromosome
sets but an addition “spindle” was created by the interaction of the back to
back astral microtubules. 4. This extra
spindle now dictates the formation of an extra cleavage furrow. Note that this results in four cells as would
have arisen without the glass rod intervention.
What is the signal from
the spindle that stimulates furrow formation?
Several candidates have been proposed to constitute this
signal. These include the protein
INCENP (Eckley et al, 1997) a chromosome passenger inner centromere protein
that is one of the very earliest (if not the earliest) components of the
cleavage furrow, before any accumulation of myosin II. The properties that such a protein would
need are to be able to recognise anti-parallel microtubules (in order to locate
the spindle pole), and some sort of regulatory activity, possibly a kinase,
that would result in the accumulation of active components of the contractile
ring?
Constituents of
the cleavage furrow.
The cleavage furrow is composed primarily
of actin arranged as concentric anti-parallel filaments. Many actin bundling proteins are known to
concentrate within the cleavage furrow, while others are excluded. It is important for the contractile function
of the furrow to have bundling proteins that bundle filaments in an
anti-parallel fashion (to allow myosin II to act on them) and to bundle with a
spacing compatible with myosin II mini-filament interaction.. Other molecules
in the furrow include G-protein Rho, Rho kinases and many myosin kinases.
Myosin II is a crucial for contraction and so is regulated by many different
kinases at different stages. When the
furrow is forming protein kinase C phosphorylates the myosin preventing its
activity. Then Cdc2 activates myosin
and the contractile activity squeezes the furrow. The furrow does not gain in thickness as it contracts and so a
net loss of actin and other constituents must take place. The cofilins are important in this activity,
and cytokinesis is impaired in cofilin mutants. The actin binding/severing activity of cofilin is regulated by
phosphorylation by LIMkinase1 and 2, but its not yet clear if either of these
kinases is involved in the regulation of cofilin at cytokinesis. In addition to myosin and actin the following
ABPs are found in the cleavage furrow:-
|
Protein |
Function |
|
Radixin |
A
member of the ERM group, radixin binds the contractile ring to the membrane. |
|
Cofilin |
Depolymerises
actin filaments as contractile ring contracts. Under control of the LIM
kinases. |
|
a-actinin |
Bundles
filaments together. Can bundle microfilaments in a parallel or anti-parallel
fashion. |
|
Talin |
Binds
the contractile ring to the membrane with radixin. |
|
Cortexillins |
Bundling
proteins from Dictyostelium with a
PIP2 binding C-terminal region important for targeting to the cleavage furrow
and cytokinesis. |
Two models have been forwarded to explain how pole-ward microtubule
flux drives the chromosomes towards the poles.
Minus end directed motor
protein in the kinetochore hydrolyses ATP to move along the attached MT Kinetochore microtubule An alternative view is that
kinetochore behaves like a thermal ratchet (compare this with Figure CS3). This
ratchet works in reverse mode, with the kinetochore “chasing” the
microtubule end.
![]()


Kinetochore microtubule

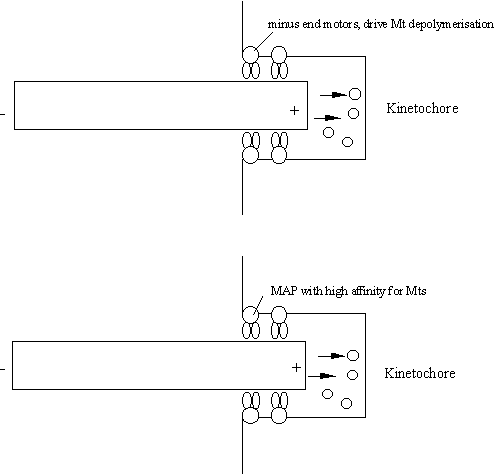

Depolymerisation causes the
kinetochore to move as it binds to the MT end every so often as it vibrated
in a Brownian fashion.
Figure 42
|
Kinetochore microtubule |
|
Depolymerisation causes the
kinetochore to move as it binds to the MT end every so often as it vibrated
in a Brownian fashion. |
An unconnected (lost) kinetochore can stop the whole process
until it is found by a microtubule and pulled back into line with the others at
the mid-zone. Then the next step,
chromosome separation can occur. How
this works is largely a mystery!
Figure 43.
The role of Myosin
II in cytokinesis
The conventional, two headed myosin, myosin
II, is involved at every stage of cytokinesis and all cytoskeletal changes that
occur before and after the actual separation process.
Separation of cells
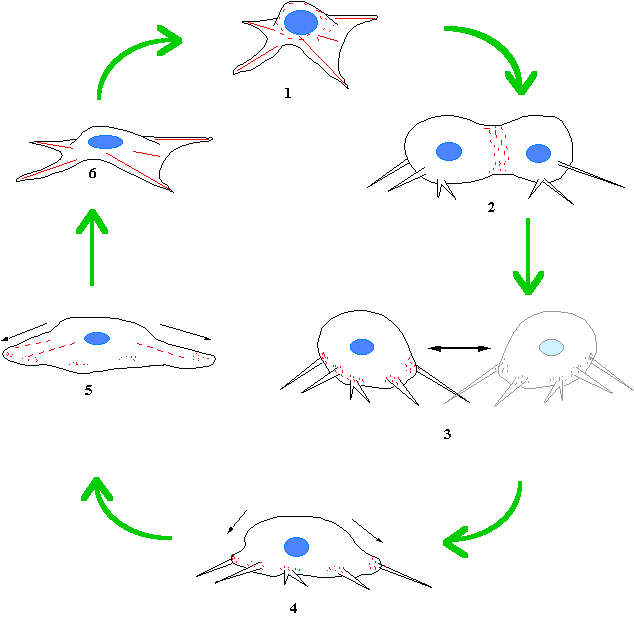
![]()
Figure 44. The role of myosin II at all stages of cytokinesis. 1. As the cell rounds up at the onset of cytokinesis, myosin
moves from the depolymerizing stress fibres into the cortex of the rounding cell.
Myosin II helps this process by loosening the stress fibres and by contraction
of the gathering cortex. 2. The main job is to contract and form the
cleavage furrow. 3. On completion of cytokinesis, the cell is
still attached to the substrate by focal adhesion rudiments through “retraction
filaments”. 4. The cell then pulls itself back to the substrate using
myosin II to work down the actin bundles within the retraction filament. Note
that the filaments in the retraction filament are polar, with the pointed ends
all facing the cell body, the correct orientation for a productive interaction
with myosin II. 5. Myosin continues to spread the cell, and
the stress fibres are re-established.
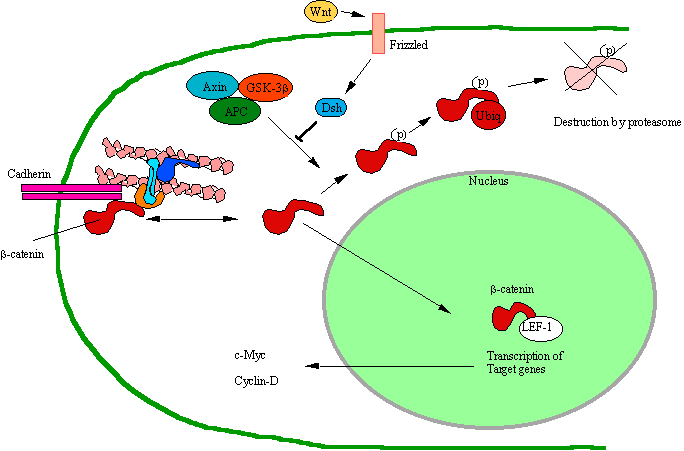
Figure 45. The Wnt Pathway. The Wnt pathway connects mitosis to the cell-cell
connectivity. This is how the cell
“knows” what shape it is in and how the shape affects the cell cycle. APC =
Adenomatous Polyposis Coli protein.
References
Heald, R. & Walczak 1999 Microtubule-based motor function in
mitosis. Curr.Op.Cell Biol. 9,
268-274.
Wolf, W.A., Chew, T.-L. & Chisolm, R.L. 1999 Regulation of
cytokinesis. Cell.Mol.Life Sci. 55,
108-120.
Rieder, C.L. & Slamon, E.D. 1998 The vertebrate cell kinetochore
and oits role during mitosis. Trends Cell Biol. 8, 310-318.
Eckley, D.M., Ainszein, A.M. Mackay, A.M., Goldberg, I.G. &
Earnshaw, W.C. 1997. Chromosome proteins and cytokinesis Patterns of cleavage
furrow formation and inner centromere positioning in mitotic heterokaryons and
mid-anaphase cells. J.Cell Biol. 136,
1169-1183.
Wheatley, S.P.1999 Updates on the mechanics and regulation of
cytokinesis in animal cells Cell Biol.Int.
23, 797-803.
Gumbiner, B.M 1997 Carcinogenesis: A balance between b-catenein and APC. Curr.Biol. 7, R443-R446.
Room 444 or lab 446 fourth floor HRB. Tel (0131) 650 3714 or 3712. E-mail SKM@srv4.med.ed.ac.uk