| EDInfo | Biomedical Sciences | Maciver Lab. Home | ABP A-Z Encyclopaedia | Amoebae | Protist Links | Cytoskeleton Links | Site Index |  |
|---|
|
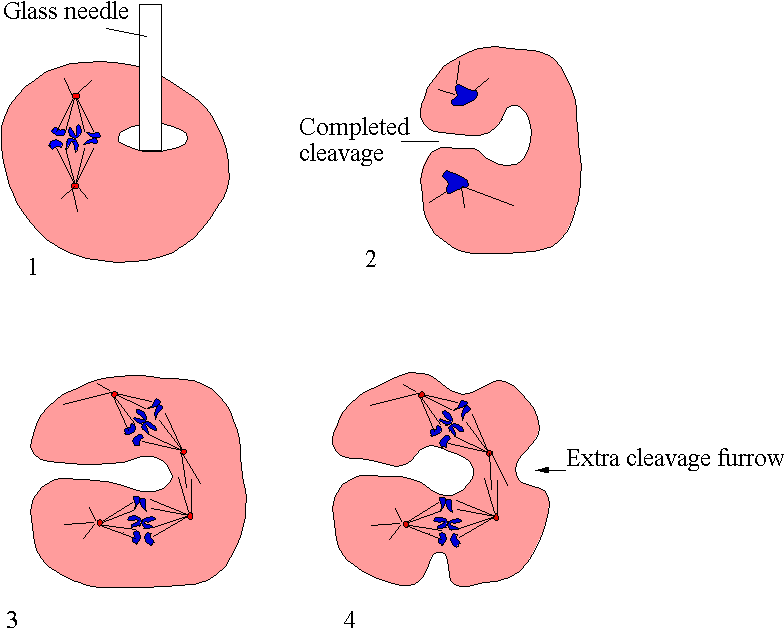
The cleavage furrow is an actin rich "purse string" that draws tight to separate daughter cells to complete cytokinesis in cell division. Early work established that the contractile ring was composed of filaments (Schroeder, 1968), which were later identified as being of actin (Perry et al, 1971; Schroeder, 1973) and myosin II (Fujiwara & Pollard, 1976). Since then a host of actin binding proteins and other cytoskeletal proteins have been identified as being constituents of the cleavage furrow. See Table 1 for a list of actin binding proteins known, or suspected of being involved in cleavage formation and function, and Table 2 for another (shorter), list of actin binding proteins for which there is evidence (usually the failure to co-localise to the cleavage furrow) against an involvement. More information on each of the individual actin-binding proteins is available from the Encyclopaedia of ABPs. It is important to note that the actin -binding proteins listed in table 1 are necessary for classic cytokinesis not traction mediated cytokinesis (see below for the distinction). In most cases these ABPs role in traction mediated cytokinesis has not been tested. Yeasts both fission (Schizosaccharomyces pombe) and budding (Saccharomyces cerevisiae) are similar enough to reveal details of animal cytokinesis. Plants do things rather differently and Plant Cytokinesis is discussed separately as is Bacterial Cytokinesis. When cytokinesis goes wrong this can result in unequal separation of chromosomes leading to genetic instability and cancer (Cytokinesis & Cancer). Asymmetric cytokinesis is important for morphogenesis in development.
There are a few inconsistencies between various cell types as to the actual requirements of some ABPs for cytokinesis, for example Dictyostelium can apparently do without a-actinin for cytokinesis (Furukawa & Fechheimer, 1994), whereas it is present in the cleavage furrow of other cell types (Fujiwara et al, 1978) where is thought to be required.
Cytokinesis Links On this website:- External links:- References:- Adachi, H., Takahashi, Y., Hasebe, T., Shirouzu, M., Yokoyama, S. & Sutoh, K. (1997) Dictyostelium IQGAP-related protein specifically involved in the completion of cytokinesis. J.Cell Biol. 137, 891-898. Adams, R. R., Wheatley, S. P., Gouldsworthy, A. M., Kandeis-Lewis, S. E., Carmena, M., Smythe, C., Gerloff, D. L. & Earnshaw, W. C. (2000) INCENP binds the Aurora-related kinase AIRK2 and is required to target it to chromosomes, the central spindle and cleavage furrow. Curr.Biol. 10, 1075-1078. Andreassen, P. R., Palmer, D. K., Wener, M. H. & Margolis, R. L. (1991) Telophase disc: a new mammalian mitotic organelle that bisects telophase cells with a possible function in cytokinesis. J. Cell Sci. 99, 523-534. Balasubramanian, M. K., Hirani, B. R., Burke, J. D. & Gould, K. L. (1994) The Schizosaccharomyces-pombe cdc3+ gene encodes a profilin essential for cytokinesis, Journal Of Cell Biology. 125, 1289-1301. Balasubramanian, M. K., McCollum, D. & Surana, U. (2000) Tying the knot: linking cytokinesis to the nuclear cycle., J.Cell Sci. 113, 1503-1513. Biron, D., Libros, P., Sagi, D., Mirelman, D. & Moses, E. (2001) 'Midwives' assist dividing amoebae., Nature. 410, 430. Bischoff, J.R., Anderson, L., Zhu, Y., Mossie, K., Ng, L., Souza, B., Schryver, B., Flanagan, P., Clairvoyant, F., Ginter, C. Chan, C.S, Novotny, M., Slamon, D.J., & Plowman, G.D. (1998). EMBO J. 17, 3052-3065. Brown, J.M., Hardin, C. & Gaertig, J. (1999). Rotokinesis, a novel phenomenon of cell locomotion-assisted cytokinesis in the ciliate Tetrahymena thermophila. Cell Biol. Int. 23, 841-848. Burgess, R.W., Deitcher, D.L., & Schwartz, T.L. (1997). The synaptic protein syntaxin 1 is required for cellularization of Drosophila embryos. J.Cell Biol. 138, 861-875. Burton, K. & Taylor, D. L. (1997) Traction forces of cytokinesis measured with optically modified elastic substratum. Nature. 385, 450-454. Bowerman, B. & Severson, A. F. (1999) Cell division: plant-like properties of animal cell cytokinesis. Current Biology. 9, R658-R660. Cao, L.-g. & Wang, Y.-L. (1990) Mechanism of the formation of contractile ring in dividing animal cells. I Recruitment of preexisting actin filaments into the cleavage furrow., J.Cell Biol. 110, 1089-1095. Cao, L.-g. & Wang, Y.-L. (1990) Mechanism of the formation of contractile ring in dividing animal cells. II Cortical movement of microinjected actin filaments., J.Cell Biol. 111, 1905-1911. Cao, L.-G. & Wang, Y.-L. (1996) Signals from the spindle midzone are required for the stimulation of cytokinesis in cultured epithelial cells, Mol.Biol.Cell. 7, 225-232. Chang, F., Drubin, D. & Nurse, P. (1997) cdc12p, a protein required for cytokinesis in fission yeast, is a component of the cell division ring and interacts with profilin., J.Cell Biol. 137, 169-182. Chalkey, H.W. (1935). The mechanism of cytoplasmic fission in Amoeba proteus. Protoplasma 24, 607-621. Chalkey, H.W. (1951). Control of fission in Amoeba proteus as related to the mechanism of cell division. Ann.N.Y. Acad.Sci. 51, 1303-1310. Cheney, R. E. & Rodriguez, O. C. (2001) A switch to release the motor, Science. 293, 1263-1264. Cheng, A., Dean, N. M. & Honkanen, R. E. (2000) Serine/threonine protein phosphatase type1g1 is required for the completion of cytokinesis in human A549 lung carcinoma cells., J.Biol.Chem. 275, 1846-1854. Conner, S.D. & Wessel, G.M. (1999). Syntaxin is required for cell division Mol.Biol.Cell 10, 2735-2743. Cooke, C.A., Heck, M.M., Earnshaw, W.C. (1987). The inner centromere protein (INCENP) antigens: movement from inner centromere to midbody during cytokinesis. J.Cell Biol. 105, 2053-2067. de Hostos, E. L., Rehfuess, C., Bradtke, B., Waddell, D. R., Albrecht, R., Murphy, J. & Gerisch, G. (1993) Dictyostelium mutants lacking the cytoskeletal protein coronin are defective in cytokinesis and cell motility, J. Cell Biol. 120, 163-173. de Lozanne, A. & Spudich, J.A. (1987). Disruption of the Dictyostelium myosin heavy-chain gene by homologous recombination. Science 236, 1086-1091. Dumontier, M., Hocht, P., Mintert, U. & Faix, J. (2000) Rac1 GTPases control filopodia formation, cell motility, endocytosis, cytokinesis and development in Dictyostelium., J.Cell Sci. 113, 2253-2265. Dutartre, H., Davoust, J., Gorvel, J.-P. & Chavrier, P. (1996) Cytokinesis arrest and redistribution of actin-cytoskeleton regulatory components in cells expressing the Rho GTPase CDC42Hs, J. Cell Sci. 109, 367-377. Eckley, D. M., Ainsztein, A. M., Mackey, A. M., Goldberg, I. & Earnshaw, W. C. (1997) Chromosomal proteins and cytokinesis: Patterns of cleavage furrow formation and inner centromere protein positioning in mitotic heterokaryons and mid-anaphase cells, J.Cell Biol. 136, 1169-1183. Edamatsu, M., Hirono, M. & Watanabe, Y. (1992) Tetrahymena profilin is localized in the division furrow, J. Biochemistry. 112, 637-642. Emoto, K., Kobayashi, T., Yamaji, A., Aizawa, H., Yahara, I., Inoue, K. & Umeda, M. (1996) Redistribution of phsophatidylethanolamine at the cleavage furrow of dividing cells during cytokinesis. PNAS 93, 12867-12872. Emoto, K. & Umeda, M. (2001) Membrane lipid control of cytokinesis. Cell Structure & Function. 26, 659-665. Eng, K., Naqvi, N. I., Wong, K. C. Y. & Balasubramanian, M. K. (1998) Rng2p, a protein required for cytokinesis in fission yeast, is a component of the actomyosin ring and the spindle pole body, Current Biol. 8, 611-621. Faix, J., Steinmetz, M., Boves, H., Kammerer, R. A., Lottspeich, F., Mintert, U., Murphy, J., Stock, A., Aebi, U. & Gerisch, G. (1996) Cortexillins, major determinants of cell shape and size, are actin bundling proteins with a parallel coiled-coil tail, Cell. 86, 631-642. Field, C. M. & Alberts, B. M. (1995) Anillin, a contractile ring protein that cycles from the nucleus to the cell cortex, J.Cell Biol. 131, 165-178. Field, C., Li, R. & Oegema, K. (1999) Cytokinesis in eukaryotes: a mechanistic comparison, Curr.Op.Cell Biol. 11, 68-80. Franck, Z., Footer, M. & Bretscher, A. (1990) Microinjection of villin into cultured cells induces rapid and long-lasting changes in cell morphology but does not inhibit cytokinesis, cell motility or membrane ruffling., J. Cell Biol. 111, 2475-2485. Fujiwara, K. & Pollard, T. D. (1976) Fluorescent antibody localization of myosin in the cytoplasm, cleavage furrow and mitotic spindle of human cells., J.Cell Biol. 71, 848-875. Fukui, Y. & Inoue, S. (1991). Cell division in Dictyostelium with special emphasis on actomyosin organization in cytokinesis. Cell Mot. Cytoskeleton 18, 41-54. Fukui, Y., Engler, S., Inoue, S. & de Hostos, E. L. (1999) Architectural dynamics and gene replacement of coronin suggests its role in cytokinesis., Cell Motility Cytoskeleton. 42, 204-217. Furukawa, R. & Fechheimer, M. (1994) Differential localization of a-actinin and the 30 kD actin-bundling protein in the cleavage furrow, phagocytic cup, and contractile vacuole of Dictyostelium discoideum, Cell Motility and Cytoskeleton. 29, 46-56. Gerisch, G. & Weber, I. (2000) Cytokinesis without myosin II., Curr.Op.Cell Biol. 12, 126-132. Giansanti, M. G., Bonaccorsi, S. & Gatti, M. (1999) The role of anillin in meiotic cytokinesis of Drosophila males., J.Cell Science. 112, 2323-2334. Glotzer, M. (1997) Cytokinesis, Curr.Biol. 7, 274-276. Golsteyn, R.M., Mundt, K.E., Fry, A.M., & Nigg, E.A. (1995) J.Cell Biol. 129, 1617-1628. Gunsalus, K. C., Bonaccorsi, S., Williams, E., Verni, F., Gatti, M. & Goldberg, M. L. (1995) Mutations in twinstar, a Drosophila gene encoding a cofilin/ADF homologue, result in defects in centrosome migration and cytokinesis, J. Cell Biol. 131, 1243-1259. Hales, K. G., Bi, E., Wu, J.-Q., Adam, J. C., Yu, I.-C. & Pringle, J. R. (1999) Cytokinesis: an emerging unified theory for eukaryotes?, Curr.Op.Cell Biol. 11, 717-725. Haugwitz, M., Noegel, A. A., Karakesisoglou, J. & Schleicher, M. (1994) Dictyostelium amoebae that lack G-actin sequestering profilins show defects in F-actin content, cytokinesis and development., Cell. 79, 303-314. Hiramoto, Y. (1975) Force exerted by the cleavage furrow of sea urchin eggs. Devel. Growth Diff. 17, 27- Hirose, K., Kawashima, T., Iwamoto, I., Nosaka, T. & Kitamura, T. (2001) MgcRacGAP is involved in cytokinesis through associating with mitotic spindle and midbody., J.Biochem. 276, 5821-5828. Insall, R. H., Muller-Taubenberger, A., Machesky, L. M., Kohler, J., Simmeth, E., Atkinson, S. J., Weber, I. & Gerisch, G. (2001) Dynamics of the Dictyostelium Arp2/3 complex in endocytosis, cytokinesis, and chemotaxis., Cell Motility Cytoskeleton. 50, 115-128. Iwasaki, T., Murata-Hori, M., Ishitobi, S. & Hosoya, H. (2001) Diphosphorylated MRLC is required for organization of stress fibers in interphase cells and the contractile ring in dividing cells. Cell Structure & Function. 26, 677-683. Jantsch-Plunger, V. & Glotzer, M. (1999) Depletion of syntaxins in the early Caenrhabditis elegans embryo reveals a role for membrane fusion events in cytokinesis., Curr. Biol. 9, 738-745. Jantsch-Plunger, V., Gonczy, P., Romano, A., Schnabel, H., Hamill, D., Schnabel, R., Hyman, A.A. & Glotzer, M. (2000) J.Cell Biol. 149, 1391-1404. Jensen, C.G. & Watson, M. (1999). Inhibition of cytokinesis by asbestos and synthetic fibres. Cell Biol. Int. 23(12), 829-840. Jiang, W., Jimenez, G., Wells, N. J., Hope, T. J., Wahl, G. M., Hunter, T. & Fukunaga, R. (1998) PRC1: A human mitotic spindle-associated CDK substrate protein required for cytokinesis., Mol.Cell. 2, 877-885. Kaitna, S., Mendoza, M., Jantsch-Plunger, V. & Glotzer, M. (2000) Incenp and an Auora-like kinase form a complex essential for chromosome segregation and efficient completion of cytokinesis., Curr.Biol. 10, 1172-1181. Karcher, R. L., Roland, J. T., Zappacosta, F., Hudddleston, M. J., Annan, R. S., Carr, S. A. & Gelfand, V. I. (2001) Cell cycle regulation of myosin-V by Calcium/Calmodulin-dependent protein kinase II., Science. 293, 1317-1320. Kawano, Y., Fukata, Y., Oshiro, N., Amano, M., Nakamura, T., Ito, M., Matsumura, F., Inagaki, M. & Kaibuchi, K. (1999) Phosphorylation of myosin-binding subunit (MBS) of myosin phosphatase by Rho-kinase during cell migration and cytokinesis., J.Cell Biol. 147, 1023-1037. Kishi, K., Sasaki, T., Kuroda, S., Itoh, T. & Takai, Y. (1993) J.Cell Biol. 120, 1187-1195. Knecht, D. A. & Loomis, W. F. (1987) Antisense RNA inactivation of myosin heavy chain gene expression in Dictyostelium discoideum., Science. 236, 1081-1085. Konzok, A., Weber, I., Simmeth, E., Hacker, U., Maniak, M. & Muller-Taubenberger, A. (1999) DAip1, a Dictyostelium homologue of the yeast actin-interacting protein 1, is involved in endocytosis, cytokinesis, and motility., J.Cell Biol. 146, 453-464. Kuczmarski, E.R. & Spudich, J.A. (1980). Regulation of myosin self-assembly: phosphorylation of Dictyostelium heavy chain inhibits formation of thick filaments. PNAS 77, 7292-7296. Larochelle, D. A., Vithalani, K. K. & DeLozanne, A. (1996) A novel member of the rho family of small GTP-binding proteins is specifically required for cytokinesis, J.Cell Biol. 133, 1321-1329. Lippincott, J. & Li, R. (1998) Sequential assembly of myosin II, an IQGAP-like protein, and filamentous actin to a ring structure involved in budding yeast cytokinesis, J.Cell Biol. 140, 355-366. Liu, T., Williams, J.G., Clarke, M. (1992). Inducible expression of calmodulin antisense RNA in Dictyostelium cells inhibits the completion of cytokinesis. Mol.Biol.Cell 3, 1403-1413. Mabuchi, I.& Okuno, M. (1977). The effect of myosin antibody on the division of starfish blastomeres. J.Cell Biol. 74, 251-263. Mabuchi, I. (1994) Cleavage furrow: timing of emergence of contractile ring actin filaments and establishment of the contractile ring by filament bundling in sea urchin eggs, J. Cell Sci. 107, 1853-1862. Mabuchi, I., Hamaguchi, Y., Fujimoto, H., Morii, N., Mishima, M. & Narumiya, S. (1993) A Rho-like protein is involved in the organization of the contractile ring in dividing sand dollar eggs. Zygote 1, 325-331. Maciver, S. K. (1996) Myosin II function in non-muscle cells, BioEssays. 18, 179-182. Madaule, P., Eda, M., Watanabe, N., Fujisawa, K., Matsuoka, T., Bito, H., Ishizaki, T., & Narumiya, S. (1998) Role of citron kinases as a target of the small GTPase rho in cytokinesis. Nature 394, 491-494. Maddox, A. S. & Burridge, K. (2003) RhoA is required for cortical retraction and rigidity during mitotic cell rounding. J.Cell Biol. 160, 255-265. Mak, A.S., Carpenter, M., Smillie, L.B., & Wang, J.H. (1991) Phosphorylation of caldesmon by p34cdc2 kinase. Identification of phosphorylation sites. J.Biol.Chem. 266, 19971-19975. Matsumura, F., Ono, S., Yamakita, Y., Totsukawa, G. & Yamashiro, S. (1998) Specific localization of serine 19 phosphorylated myosin II during cell locomotion and mitosis of cultured cells., J.Cell Biol. 140, 119-129. Maupin. P. & Pollard, T.D. (1986). Arrangement of actin filaments and myosin-like filaments in the contractile ring and of actin-like filaments in the mitotic spindle of dividing HeLa cells. J.Ultrastructure Mol. Struct.Res. 94, 92-103. Megraw, T. L., Kao, L.-R. & Kaufman, T. C. (2001) Zygotic development without functional mitotic centrosomes., Curr.Biol. 11, 116-1120. Miller, A. L., Fluick, R. A., McLaughlin, J. A. & Jaffe, L. F. (1993) Calcium buffer injections inhibit cytokinesis in Xenopus eggs, J. Cell Sci. 106, 523-534. Mitchison, T. J. (2001) Psycosine, cytokinesis, and orphan receptors: Unexpected connections. J.Cell Biol. 153, F1-F3. Mullins, J.M. & Biesele, J.J. (1977). Terminal phase of cytokinesis in D-98S cells. J.Cell Biol. 73, 672-684. Nagaoka, R., Abe, H., Kusano, K. & Obinata, T. (1995) Concentration of cofilin, a small actin-binding protein, at the cleavage furrow during cytokinesis, Cell Motility And The Cytoskeleton. 30, 1-7. Nagasaki, A., Hibi, M., Asano, Y. & Uyeda, T. Q. P. (2001) Genetic approaches to dissect the mechanisms of two distinct pathways of cell cycle-coupled cytokinesis in Dictyostelium. Cell Structure & Function. 26, 585-591. Niewöhner, J., Weber, I., Maniak, M., Müller-Taubenberger, A. & Gerisch, G. (1997) Talin-null cells of Dictyostelium are strongly defective in adhesion to particle and substrate surfaces and slightly impaired in cytokinesis, J.Cell Biol. 138, 349-361. Nigg, E.A. (1998) Polo-like kinases: positive regulators of cell division from start to finnish. Curr.Op.Cell Biol. 10, 776-783. Nislow, C., Lombillo, V.A., Kuriyama, R. & McIntosh, J.R. (1992). A plus-end-directed motor enzyme that moves antiparallel microtubules in vitro localizes to the interzone of mitotic spindles. Nature 359, 543-547. Noegel, A. A., Rivero, F., Albrecht, R., Janssen, K.-P., Kohler, J., Parent, C. A. & Schleicher, M. (1999) Assessing the role of the ASP56/CAP homologue of Dictyostelium discoideum and the requirements for subcellular localization., J.Cell Sci. 112, 3195-3203. Noguchi, T. & Mabuchi, I. (2000) Reorganization of actin cytoskeleton at the growing end of the cleavage furrow of Xenopus egg during cytokinesis., J.Cell.Sci. 114, 401-412. O'Connel, C.B., Wheatley, S.P., Ahmed, S. & Wang, Y.L. (1999). The small GTP-binding protein Rho regulates cortical activites in culture cells during division. J.Cell Biol. 144, 305-313. Oh, S. W. & Jeon, K. W. (1998) Characterization of myosin heavy chain and its gene in Amoeba proteus, J.Euk.Microbiol. 45, 600-605. Perry, M.M., John, H.A., Thomas, N.S.T. (1971) Actin-like filaments in the cleavage furrow of newt eggs. Exp.Cell Res. 65, 249. Pollard, T.D., Satterwhite, L., Cisek, L., Corden, J., Sato, M., & Maupin, P. (1990). Actin and myosin in relation to cytokinesis. In "Cytokinesis: Mechanisms of furrow formation during cell division." Ann.N.Y.Acad.Sci. 582, 120-130. Rappaport, R. (1967). Cell division. Direct measurement of maximum tension exerted by furrow of echinoderm eggs. Science 156, 1241-1243. Rappaport, R. (1986). Establishment of the mechanism of cytokinesis in animal cells. Int.Rev.Cytol. 105, 245-281. Rivero, F., Furukawa, R., Noegel, A. A. & Fechheimer, M. (1996) Dictyostelium discoideum cells lacking teh 34,000-dalton actin binding protein can grow, locomote, and develop, but exhibit defects in regulation of cell structure and movement: A case of partial redundancy, J.Cell Biol. 135, 965-980. Rivero, F., Furukawa, R., Fechheimer, M. & Noegel, A. A. (1999) Three actin cross-linking proteins, the 34 kDa actin bundling protein, a-actinin and gelation factor (ABP-120), have both unique and redundant roles in the growth and development of Dictyostelium., J.Cell Sci. 112, 2737-2751. Robinson, D. N. & Spudich, J. A. (2000) Dynacortin, a genetic link between equatorial contractility and global shape control discovered by library complementation of a Dictyostelium discoideum cytokinesis mutant., J.Cell Biol. 150, 823-838. Robinson, D. N., Cavet, G., Warrick, H. M. & Spudich, J. A. (2002) Quantitation of the distribution and flux of myosin-II during cytokinesis. BMC Cell Biology. 3, 1-13. Sabry, J. H., Moores, S. L., Ryan, S., Zang, J.-H. & Spudich, J. A. (1997) Myosin heavy chain phosphorylation sites regulate myosin localization during cytokinesis in live cells., Mol. Biol.Cell. 8, 2605-2615. Sanger, J. M., Dome, J. S., Hock, R. S., Mittal, B. & Sanger, J. W. (1994) Occurence of fibers and their association with talin in the cleavage furrows of PtK2 cells, Cell Mot.Cytoskeleton. 27, 26-40. Sarto, N., Yonemura, S., Obinata, T., Tsukita, S. & Tsukita, S. (1991) Radixin, a barbed end capping actin-modulating protein is concentrated at the cleavage furrow during cytokinesis, J. Cell Biol. 113, 321--330. Satterwhite, L. S. & Pollard, T. D. (1992) Cytokinesis, Current Biol. 4, 43-52. Satterwhite, L. S. , Lohka, M.J., Wilson, K.L., Scherson, T.Y., Cisek, L.J., Corden, J.L., & Pollard, T. D. (1992) Phosphorylation of myosin-II regulator light chain by cyclin-p34cdc2: a mechanism for the timing of cytokinesis. J.Cell Biol. 118, 595-605. Saunders, W.S., Shuster, M., Huang, X., Gharaibeh, B., Enyenihi, A.H., Petersen, I., & Gollin, S.M. (2000). Chromosomal instability and cytoskeletal defects in oral cancer cells. PNAS 97, 303-308 Schroeder, T.E. (1968). Cytokinesis: filaments in the cleavage furrow. Exp.Cell Res. 53, 272. Schroeder, T.E. (1972). Actin in dividing cells: contractile ring filaments bind heavy meromyosin. PNAS 70, 1688-1692. Schroeder, T.E. (1976). Actin in dividing cells: Evidence for its role in cleavage but not mitosis. "Cell Motility". Cold Spring Harbor Conferences on Cell Proliferation. 3, 265-277 Severson, A. F., Hamill, D. R., Carter, J. C., Schumacher, J. & Bowerman, B. (2000) The Aurora-related kinase AIR-2 recruits ZEN-4/CeMKLP1 to the mitotic spindle at metaphase and is required for cytokinesis., Curr.Biol. 10, 1162-1171. Shapiro, P.S., Vaisberg, E., Hunt, A.J., Tolwinski, N.S., Whalen, A.M., McIntosh, J.R., & Ahn, N.G. (1998) J.Cell Biol. 142, 1533-1545. Shu, S., Lee, R.J., LeBlanc-Straceski, J.-M. & Uyeda, T.Q.P. (1999). Role of myosin II tail sequences in its function and localization at the cleavage furrow in Dictyostelium. J.Cell Sci. 112, 2195-2201. Shuster, C. B. & Burgess, D. R. (1999) Parameters that specify the timing of cytokinesis, J.Cell Biol. 146, 981-992. Smith, J.L., Silveira, L.A. & Spudich, J.A. (1996). Myosin light chain kinase (MLCK) gene disruption in Dictyostelium: a role for MLCK-A in cytokinesis and evidence for multiple MLCKs PNAS 93, 12321-12326 Stock, A., Steinmertz, M. O., Janmey, P. A., Aebi, U., Gerisch, G., Kammerer, R. A., Weber, I. & Faix, J. (1999) Domain analysis of cortexillin I: actin bundling, PIP2-binding and the rescue of cytokinesis., EMBO. 18, 5274-5284. Straight, A. F. & Field, C. M. (2000) Microtubules, membranes and cytokinesis, Curr.Biol. 10, R760-R770. Suetsuga, S., Miki, H. & Takenawa, T. (1999) Distinct roles of profilin in cell morphogenesis changes: microspikes, membrane ruffles, stress fibers, and cytokinesis., FEBS. tba. Swan, K.A., Severson, AF., Carter, J.C., Martin, P.R., Schabel, H., Schnabel, R. & Bowerman, B. (1998). Cyk-1: a C.elegans FH gene required for a late step in embryonic cytokinesis. J.Cell Sci. 111, 2017-2027. Tatsumoto, T., Xie, X., Blumenthal, R., Okamoto, I. & Miki, T. (1999) Human ECT2 is an exchange factor for Rho GTPases, phosphorylated in G2/M phases, and involved in cytokinesis., J.Cell Biol. 147, 921-927. Takaishi, K., Sasaki, T., Kameyama, T., Tsukita, S. & Takai,Y. (1995) Oncogene 11, 39-48. Terada, Y., Tatsuka, M., Suzuki, F., Yasuda, Y., Fujita, S., & Otsu, M. (1998) Aim-1: a mammalian midbody-associated protein required for cytokinesis. EMBO J. 17, 667-676. Theriot, J. A. & Satterwhite, L. L. (1997) New wrinkles in cytokinesis, Nature. 385, 388-389. Tosuji, H., Mabuchi, I., Fusetani, N. & Nakazawa, T. (1992) Calyculin A induces contractile ring-like apparatus formation and condensation of chromosomes in unfertilized sea urchin eggs, Cell Biol. 89, 10613-10617. Tuxworth, R. I., Cheetham, J. L., Machesky, L. M., Spiegelmann, G. B., Weeks, G. & Insall, R. H. (1997) Dictyostelium RasG is required for normal motility and cytokinesis, but not growth, J.Cell Biol. 138, 605-614. Walker, G. R., Shuster, C. B. & Burgess, D. R. (1997) Microtubule-entrained kinase activites associated with the cortical cytoskeleton during cytokinesis, J.Cell Sci. 110, 1373-1386. Weber, I., Gerisch, G., Heizer, C., Murphy, J., Badelt, K., Stock, A., Schwartz, J.-M. & Faix, J. (1999) Cytokinesis mediated through the recruitment of cortexillins into the cleavage furrow, EMBO J. 18, 586-954. Weber, I. (2001) On the mechanism of cleavage furrow ingression in Dictyostelium. Cell Structure & Function. 26, 577-584. Wheatley, S. P. & Wang, Y.-l. (1996) Midzone microtubule bundles are continuously required for cytokinesis in cultured epithelial cells., J.Cell Biol. 135, 981-989. Wheatley, S. P., Hinchcliffe, E. H., Glotzer, M., Hyman, A. A., Sluder, G. & Wang, Y.-L. (1997) CDK1 inactivation regulates anaphase spindle dynamics and cytokinesis in vivo., J.Cell Biol. 138, 385-393. Wheatley, S. P. (1999) Updates on the mechanics and regulation of cytokinesis in animal cells., Cell Biol.Internat. 23, 797-803. Win, T. Z., Gachet, Y., Mulvihill, D. P., May, K. M. & Hyams, J. S. (2000) Two type V myosins with non-overlapping functions in the fission yeast Schizosaccharomyces pombe: Myo52 is concerned with growth polarity and cytokinesis, Myo51 is a component of the cytokinetic actin ring., J.Cell Sci. 114, 69-79. Wolf, W. A., Chew, T.-L. & Chisholm, R. L. (1999) Regulation of cytokinesis, Cell Mol. Life Sci. 55, 108-120. Yamakita, Y., Yamashiro, S. and Matsumura, F. (1994). In vivo phosphorylation of regulatory light chain of myosin II during mitosis of cultured cells. J.Cell Biol.124, 129-137. Yumura, S. & Uyeda, T.Q.P. (1997). Transport of myosin II to the equatorial region without its own motor activity in mitotic Dictyostelium cells. Mol. Biol. Cell 8, 2089-2099. Yumura, S. & Fukui, Y. (1998) Spatiotemporal dynamics of actin concentration during cytokinesis and locomotion in Dictyostelium., J.Cell Sci. 111, 2097-2108. Zang, J.-H., Cavet, G., Sabry, J.H., Wagner, P., Moores, S.L. & Spudich, J.A. (1997). On the role of myosin-II in cytokinesis: division of Dictyostelium cells under adhesive and non-adhesive conditions. Mol.Biol.Cell 8, 2617-2629. Zang, J.-H. & Spudich, J.A. (1998) Myosin II localization during cytokinesis occurs by a mechanism that does not require its motor domain. PNAS 95, 13652-13657. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|


{kind=link}